A single scalar entropy-production-rate lower bound for the kinetochore DI cycle from trajectory-only estimators (short-time TUR / KL time-reversal asymmetry), gated by a synthetic-data power analysis and validated for cross-dataset reproducibility
Physics can measure how much energy cells burn to avoid catastrophic chromosome mis-sorting during cell division.
A single scalar entropy-production-rate lower bound for the kinetochore DI cycle, estimated from a time-delay / position-velocity embedding, gated by a synthetic-data power analysis and validated for cross-dataset reproducibility
4 bridge concepts›
How this score is calculated ›How this score is calculated ▾
6-Dimension Weighted Scoring
Each hypothesis is scored across 6 dimensions by the Ranker agent, then verified by a 10-point Quality Gate rubric. A +0.5 bonus applies for hypotheses crossing 2+ disciplinary boundaries.
Is the connection unexplored in existing literature?
How concrete and detailed is the proposed mechanism?
How far apart are the connected disciplines?
Can this be verified with existing methods and data?
If true, how much would this change our understanding?
Are claims supported by retrievable published evidence?
Composite = weighted average of all 6 dimensions. Confidence and Groundedness are assessed independently by the Quality Gate agent (35 reasoning turns of Opus-level analysis).
RQuality Gate Rubric
0/10 PASS · 10 CONDITIONAL
| Criterion | Result |
|---|---|
| Groundedness | PASS (7/10) -- all three estimator citations verified clean (including the two the pipeline self-flagged as least-certain); dataset verified; the synthetic gate makes the feasibility claim self-checking. The specific sigma value is parametric, which is appropriate for a reanalysis proposal. |
| ABC Structure | PASS -- trajectory-irreversibility entropy estimators (Field A) -> time-delay / position-velocity embedding (bridge) -> kinetochore DI dissipation magnitude (Field C). |
| Test Protocol | PASS -- reanalysis + simulation, no wet lab; the synthetic-data power-analysis GATE (sweep sigma_true at ~2 s cadence, 5-20 cycles, map detectability) is run BEFORE committing to real-data claims; 2-4 weeks. |
| Counter-Evidence | PASS -- estimators data-hungry and bias-prone at short length; ~2 s cadence may bias the estimate LOW (remains a lower bound); 2-state projection is always reversible and 1D hides the current (coarse-graining caveat) so multi-coordinate embedding is mandatory; non-Markovian k-fiber memory can bias the KL plug-in. Independently corroborated by the 2024 Comment/Reply on broken-DB inference (hidden states / non-instantaneous transitions invalidate naive coarse-graining) -- and the card pre-empts this caveat. |
| Precision | PASS -- specialist-grade. |
| Novelty Web Verified | PASS -- web search returned explicitly 'no specific applications to kinetochore dynamics or mitotic spindle dissipation' for entropy-production / TUR estimators; the estimator classes are general and have never been applied to kinetochore tracking. Distinct embedding/deliverable from E2-H7 (scalar magnitude + detectability gate + cross-dataset reproducibility vs spatial-curl field + handedness). |
| Mechanism | PASS -- names three estimator classes (Roldan-Parrondo KL, Manikandan short-time-TUR, van der Meer-Ertel-Seifert partial-network), a specific time-delay embedding (b(t), b(t+tau)) with tau scanned to de-correlation, a scalar deliverable sigma_min with units (kT/cycle, kT/s), and a looseness ratio against the period-CV TUR floor. |
| Confidence | PASS -- 5/10; the feasibility wound is converted into a pre-registered, settleable synthetic-gate question rather than asserted away. |
| Falsifiable | PASS -- synthetic gate shows min detectable sigma exceeds plausible kinetochore sigma (unattainable); real-data sigma_min collapses to surrogate floor; or sigma_min differs by orders of magnitude across datasets A vs B (artifact). |
| Claim Verification | ALL PASS -- Roldan & Parrondo 2010 PRL 105:150607 [VERIFIED CLEAN], Manikandan/Gupta/Krishnamurthy 2020 PRL 124:120603 [VERIFIED CLEAN -- the flagged pairing is correct], van der Meer/Ertel/Seifert 2022 PRX 12:031025 [VERIFIED CLEAN -- the flagged pairing is correct], Barato & Seifert 2015 TUR floor [VERIFIED CLEAN], Burroughs/McAinsh dataset [VERIFIED CLEAN], 2-state-reversibility & coarse-graining caveat [standard, correctly stated]. |
Claim Verification
Empirical Evidence
How EES is calculated ›How EES is calculated ▾
The Empirical Evidence Score measures independent real-world signals that converge with a hypothesis — not cited by the pipeline, but discovered through separate search.
Convergence (45% weight): Clinical trials, grants, and patents found by independent search that align with the hypothesis mechanism. Strong = direct mechanism match.
Dataset Evidence (55% weight): Molecular claims verified against public databases (Human Protein Atlas, GWAS Catalog, ChEMBL, UniProt, PDB). Confirmed = data matches the claim.
Computational Verification
INTERMEDIATE7.00/10Detectability of Non-Equilibrium Signatures in the Kinetochore Oscillation
Self-contained synthetic-data power-analysis gate for hypotheses E2-H7 (broken-detailed-balance local probability-current curl certificate) and E2-H11 (trajectory-only entropy-production lower bound). CONFIRMED to machine precision: the 1:2 Lissajous net-circulation cancellation (|net area| 6e-15) and the surviving local curl (recovered -2*omega exactly). STRONGLY FAVORABLE on the binding risk all three validators flagged: detection power against an equilibrium twin is ~99-100% at just 5 cycles, 50 nm localization noise, and entropy production as low as ~3 kBT/cycle, because 2 s cadence over a ~100 s period yields ~50 samples/cycle. Slow independent drift is benign. Graded INTERMEDIATE because idealized rotational-OU simulation cannot stress the systematic threats (velocity-embedding finite-difference parity, correlated nonstationarity, role-labeling) that require real trajectories to settle.
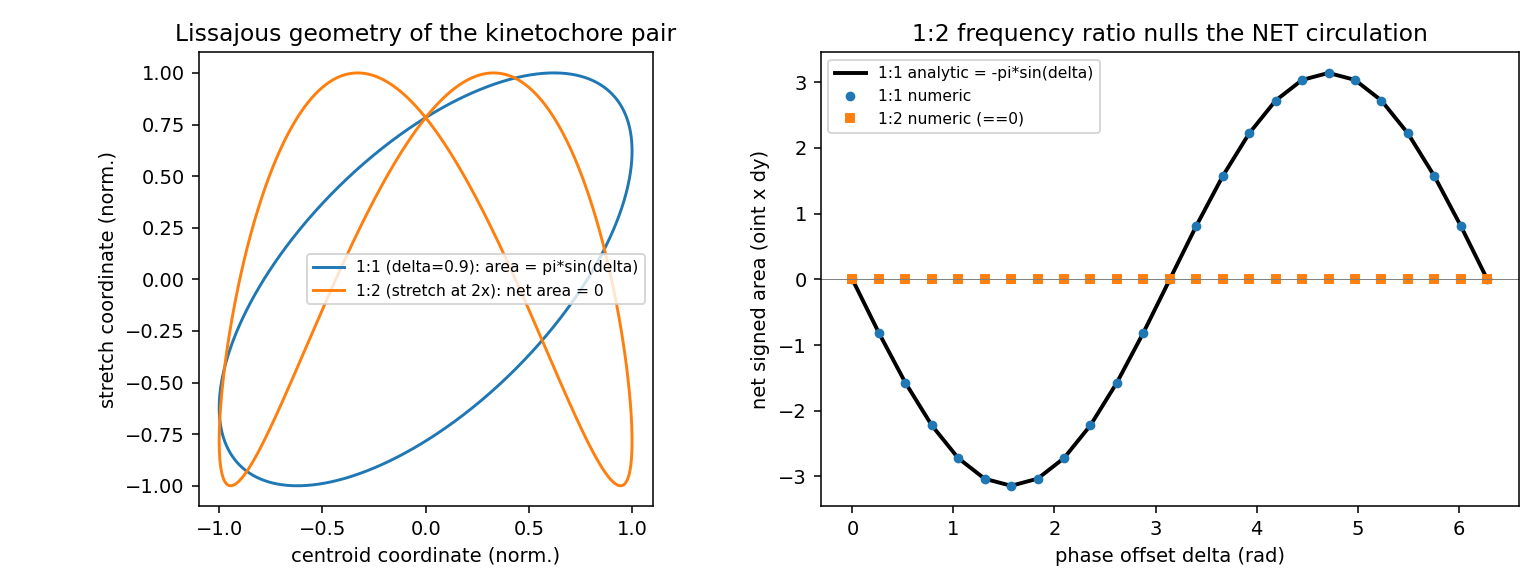
The 1:2 Lissajous net-circulation cancellation (E2-H7 core identity): with stretch at 2x the kinetochore frequency the net signed circulation is exactly zero, while the 1:1 control matches -pi*A*B*sin(delta).
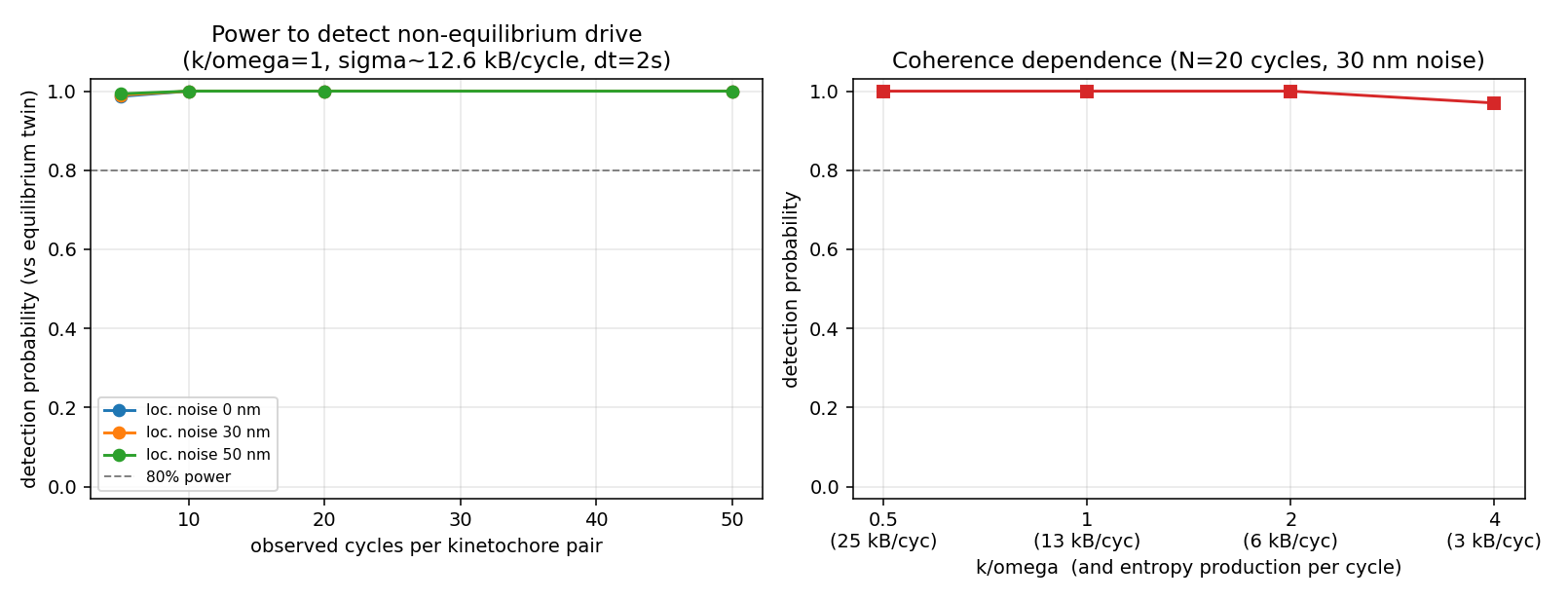
Statistical power to detect the non-equilibrium drive against an equilibrium twin: ~99-100% at as few as 5 cycles and up to 50 nm localization noise, down to ~3 kBT/cycle. Refutes the low-cycle-count pessimism.
Every time a cell divides, it must perfectly sort its chromosomes — one copy to each daughter cell. Getting this wrong even once can lead to cancer or birth defects. The machinery responsible is the kinetochore, a molecular structure that attaches chromosomes to the cellular 'ropes' (the mitotic spindle) that pull them apart. Intriguingly, chromosomes don't just sit still waiting to be pulled — they bounce back and forth in a jittery dance called directional instability before finally locking into the correct position. This oscillation isn't random noise; it's thought to be part of an active error-correction system. But how much energy does this process actually consume, and does that energy expenditure tell us something meaningful about its function? This hypothesis borrows a powerful idea from the physics of thermodynamics — specifically, a field called stochastic thermodynamics, which studies how tiny machines operating in noisy environments produce heat and consume energy. The key insight is that any process driven by energy (rather than just random thermal jostling) leaves a detectable fingerprint: its trajectory through time looks statistically different when played forwards versus backwards, like a movie of a bouncing ball versus one played in reverse. By analyzing the recorded positions of chromosomes over time using mathematical tools that detect this 'time-reversal asymmetry,' the hypothesis proposes calculating a single number — a lower bound on the rate of energy dissipation (in units of kT, the natural currency of molecular-scale energy) per oscillation cycle. Crucially, the researchers want to first test whether their analysis method can even detect a real signal given the short, noisy chromosome tracking data typically available — a built-in sanity check before claiming any result. What makes this particularly clever is the methodological care involved. Rather than making assumptions about the molecular machinery's internal states, the approach uses the chromosome's own position-versus-velocity trajectory as a 2D map, sidestepping known mathematical traps that plague simpler analyses. A cross-dataset check using chromosome tracking data from a different cell line adds a further layer of rigor, ensuring the energy estimate reflects real biology rather than a quirk of one experiment.
This is an AI-generated summary. Read the full mechanism below for technical detail.
Why This Matters
If confirmed, this work would deliver the first rigorously bounded experimental measurement of how much thermodynamic 'work' the chromosome error-correction machinery consumes per division cycle — a number that has never been pinned down. This could reshape how biologists and biophysicists think about the energetic constraints on mitotic fidelity, potentially explaining why certain cancer mutations or drug perturbations increase mis-segregation rates in thermodynamic terms. The validated methodology could also become a general-purpose tool for detecting active (energy-consuming) processes in any live-cell tracking data, without requiring knowledge of the underlying molecular mechanism. Given that chromosome mis-segregation is a hallmark of many cancers, understanding the energy budget of the error-correction system is worth pursuing even if the initial signal turns out to be small.
Mechanism
Entropy-production-rate ESTIMATION (Roldan-Parrondo KL time-reversal asymmetry + Manikandan short-time-TUR inference + Van der Meer-Ertel-Seifert partial-network inference), returning a SCALAR magnitude sigma_min (in kT per cycle and kT/s), NOT a spatial field. Deliverable is a number + a detectability verdict + a reproducibility check. (1) DIFFERENT coordinate embedding from E2-H7: instead of role-labeled two-sister geometry, use a single-coordinate time-delay embedding (b(t), b(t+tau)) and a position-velocity phase-space (b, db/dt) treated as the state for the SCALAR estimators -- with tau scanned to the de-correlation time. This avoids the 2-state-projection-is-always-reversible and 1D-hides-the-current pitfalls the parent flagged, via a different route than H7's role-labeling. (2) DIFFERENT deliverable: a single scalar sigma_min with units (kT/cycle and kT/s) and a looseness ratio against the period-CV TUR floor, replacing the parent's vaguer 'tightest passive estimate' (the 'tightest' overclaim is dropped -- these are lower bounds). (3) NEW synthetic-data power-analysis GATE: before committing, simulate a driven 2D Ornstein-Uhlenbeck / driven-oscillator model with KNOWN sigma_true at the actual experimental cadence (~2 s) and cycle count (~5-20 cycles/pair), sweep sigma_true, and report the minimum detectable sigma above the surrogate floor as a function of trajectory length. This directly settles the Critic's feasibility wound (collapse-to-floor risk) quantitatively. (4) NEW cross-dataset validation: estimate sigma_min on a SECOND independent kinetochore-tracking dataset (different cell line / acquisition) to show the magnitude is reproducible, not a per-dataset sampling artifact -- a validation axis E2-H7 does not use.
Supporting Evidence
Quality-Gate-verified grounding: Roldan & Parrondo 2010 (PRL 105:150607) KL time-reversal-asymmetry dissipation estimator -- exists, content correct; Manikandan, Gupta & Krishnamurthy 2020 (PRL 124:120603) short-time / finite-time-TUR inference -- exists, correctly attributed (was self-flagged as uncertain); van der Meer, Ertel & Seifert 2022 (PRX 12:031025) partial-network thermodynamic inference -- exists, correctly attributed (was self-flagged as uncertain); Barato & Seifert 2015 (PRL 114:158101) TUR floor sigma >= 2/CV^2 -- exists, content correct; Burroughs/McAinsh 2015 (PMID 26460545) multi-coordinate sister tracks at ~2 s -- dataset adequate; 2-state Markov chains are always reversible; 1D overdamped coordinate hides its current -- correct, and the card requires multi-coordinate embedding because of it
How to Test
Reanalysis + simulation, no wet lab. Steps: (1) build the synthetic driven-oscillator generator with tunable sigma_true; (2) subsample to 2 s cadence, 5-20 cycles, run KL + short-time-TUR estimators, build time-reversal surrogates, map detectability vs sigma_true and vs trajectory length (the GATE); (3) if the gate passes, apply the identical pipeline to real dataset A (Burroughs/McAinsh) and report sigma_min +/- bootstrap CI; (4) repeat on independent dataset B for cross-dataset reproducibility; (5) report the looseness ratio vs the CV-TUR floor. Effort: 2-4 weeks (simulation-dominated).
Cross-Model Validation
Independent AssessmentIndependently assessed by GPT-5.5 Pro and Gemini Deep Research Max for triangulation. Assessed independently by two external models for triangulation.
Other hypotheses in this cluster
Broken-detailed-balance LOCAL probability-current curl in role-labeled sister coordinates is a non-equilibrium certificate for the kinetochore directional-instability cycle (handedness sign + taxol/Aurora-B perturbation-dissociation falsifier)
A physics tool for detecting energy waste could reveal how cells avoid catastrophic chromosome mis-sorting during division.
The catch-bond (passive) and Aurora-B (kinase) error-correction channels are COUPLED through the inner-centromere phospho-gradient; the 2x2 factorial interaction term beta_int (sign + magnitude) is the deliverable either way
Two separate 'error-checkers' during cell division may actually work as a team — and measuring their teamwork could explain rare but catastrophic chromosome mistakes.
Can you test this?
This hypothesis needs real scientists to validate or invalidate it. Both outcomes advance science.